Homo Sapiens
21:31
Resumen del tema
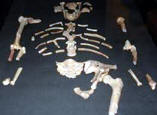
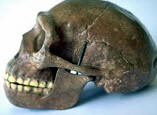
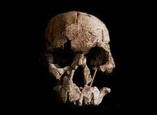

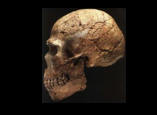
Se han propuesto dos modelos para explicar el origen de los humanos anatómicamente modernos: el primero de ellos es el modelo multirregional o "en candelabro", y el segundo es el modelo del "Arca de Noé", del origen único o de la sustitución. Según el primero, las antiguas poblaciones del Asia, África y Europa mantienen una continuidad evolutiva y genética con la humanidad actual, y son la causa de las diferencias que existen entre los diferentes grupos humanos actuales. El segundo modelo postula un origen único en un lugar geográfico concreto para la humanidad actual, y a partir de ese punto de origen una migración al resto del mundo que reemplazó a las poblaciones anteriores. El análisis de esta cuestión puede abordarse utilizando dos fuentes de información complementarias: 1) el estudio de los fósiles humanos, su taxonomía, cronología y ubicación geográfica; 2) los estudios genéticos de la variabilidad de la humanidad actual y, más excepcionalmente, de los genes que se encuentran en los fósiles. Hasta hace poco tiempo, en el estudio del registro fósil había un problema metodológico porque las técnicas de datación existentes no permitían datar los fósiles humanos de este periodo. Pero la aplicación de las técnicas de termoluminiscencia, series de uranio y resonancia de espín electrónico a permitido obtener fechas para estos restos. Estas técnicas han confirmado que el hombre moderno estaba ya presente en África hace unos 150.000-100.00 años, cuando los neandertales todavía vivían en Europa y el Homo erectus todavía poblaba la isla de Java. En Europa los hombres de Cromañón, asociados a una industria delPaleolítico superior , sustituyen bruscamente a los neandertales asociados a una industria del Paleolítico medio (musteriense ) hace unos 30.000-40.000 años. En cambio, los humanos modernos vivieron en el Próximo oriente hace unos 100.000 años, y son más antiguos que la mayor parte de los restos de neandertales hallados en este mismo lugar, por lo tanto difícilmente los primeros pueden ser descendientes de los segundos. Ambos grupos humanos presentes en Próximo Oriente compartían el mismo tipo de industria lítica, el musteriense. El esqueleto neandertal de Tabun, posiblemente, atestigua un primer poblamiento de los neandertales hace unos 110.000-100.000 años. Con posterioridad, alrededor de los 100.000 años, sólo encontramos yacimientos con fósiles de humanos modernos. Los neandertales habrían poblado el Próximo Oriente en un momento de frío intenso, que les habría obligado a desplazarse hacia el sur entre hace unos 60.000-40.000 años. Finalmente, hace unos 40.000 años, los neandertales desaparecen y sólo perviven los humanos modernos.
La variabilidad genética observada a nivel mundial es muy escasa, pero es relativamente mayor en las poblaciones africanas. La mayor parte de estudios genéticos (ADN mitocondrial, análisis cromosómico, ADN nuclear, microsatélites) coinciden en que todos los humanos modernos tenemos nuestro origen en África entre hace 200.000 y 150.000 años.Además, los análisis del ADN mitocondrial (ADNmt) de un neandertal han reforzado la opinión de que los neandertales no son antepasados de la humanidad actual, sino que ambos grupos comparten un antepasado común, lejano en el tiempo, y que podría corresponder con la especie hallada en el nivel TD6 de la Gran Dolina de la Sierra de Atapuerca: Homo antecessor. Uno de los principales problemas de los estudios genéticos es la forma de establecer los "relojes moleculares", que sitúan temporalmente los momentos de divergencia de las líneas evolutivas. Se deben tomar previamente algunos presupuestos: las semejanzas genéticas entre individuos han de ser función de su parentesco, y directamente proporcionales al tiempo de divergencia desde un antepasado común; la tasa de mutación ha de ser constante; y los genes estudiados deben ser neutros, es decir, que no actúe sobre ellos la selección natural.
Hipótesis evolutivas Desde los inicios de los estudios paleoantropológicos el origen de nuestra especie , Homo sapiens, es uno de los temas centrales de interés y debate. En primer lugar, las interpretaciones e hipótesis surgieron a partir de los fósiles humanos encontrados y de las evidencias culturales (como la industria lítica) hallados en los yacimientos arqueológicos y paleontológicos. Utilizando los mismos fósiles humanos se propusieron diferentes hipótesis evolutivas para explicar la aparición de la humanidad actual. Pero en los últimos tiempos, los estudios de tipo genético han añadido sus esfuerzos a la investigación del origen de la humanidad actual, pero, salvo en casos excepcionales que trataremos más adelante, la Genética sólo dispone del material molecular de las poblaciones actuales. Los dos tipos de información se complementan y apoyan para descubrir la forma, el cuándo y el dónde de la aparición de nuestra especie. Existen dos modelos contrapuestos para explicar el origen de los humanos anatómicamente modernos: el primero de ellos es el modelo multirregional o "en candelabro" y el segundo es el modelo del "Arca de Noé", del origen único o de la sustitución.
La hipótesis multirregional
La hipótesis de la evolución multirregional , también denominada evolución en candelabro por William Howells, contempla el proceso de aparición de nuestra especie como el resultado de una profunda transformación filética, a partir de las poblaciones ancestrales de Homo erectus en el Viejo Mundo, que evolucionaron gradual e independientemente hacia Homo sapiens arcaicos y posteriormente hasta los humanos modernos. Las diferencias que se encuentran en los diferentes continentes entre las razas actuales tendrían un origen muy antiguo y serían el resultado de un proceso evolutivo en paralelo.Esta hipótesis fue propuesta por Franz Weidenreich antes de la Segunda Guerra Mundial y defendida a mediados de siglo por Carleton Coon y otros científicos. Según Weidenreich, invocando una forma de ortogénesis, existe una dirección predeterminada en las especies para evolucionar en un determinado sentido, y que explica porque distintos linajes humanos han conducido de forma independiente a la aparición de la misma especie en diferentes lugares. Para Coon, en primer lugar habrían surgido las diferencias raciales y posteriormente habría aparecido la especie Homo sapiens. Sin embargo, a partir de los mecanismos evolutivos que conocemos, es biológicamente inviable que poblaciones aisladas genéticamente evolucionen dando lugar a una misma especie.
Por este motivo, más recientemente, los defensores del modelo mutirregional, entre los que destacan Milford Wolpoff y Alan Thorne, reconocen la importancia del flujo genético entre las distintas poblaciones geográficas. Opinan que éstas evolucionaron hacia la humanidad actual no independiente, sino de forma conjunta, con flujos genéticos entre las distintas poblaciones que formaban una compleja red genética. El flujo genético habría sido lo suficientemente importante para que las poblaciones fuesen en una misma dirección pero mantuvieron las diferencias que caracterizan a los humanos de cada región. Estos autores observan una continuidad morfológica entre los aborígenes australianos, los fósiles de Ngandong y los primeros pobladores de Java, representados por Sangiran 17. Debajo de esta hipótesis subyace una concepción de la evolución humana como una escalera ascendente que conduce inevitablemente al Homo sapiens. Es decir, nuestra especie es resultado final de la evolución a través de una serie de estadios humanos intermedios que se suceden progresivamente.
Pero como hemos podido ver a lo largo de todo este curso, los mecanismos biológicos que afectan a los humanos son los mismos que los de otros grupos animales, y la dinámica de la evolución humana no es diferente a la de otros seres vivos. El árbol evolutivo de los homínidos no es de tipo secuencial, sino que tiene una forma muy ramificada con múltiples eventos de aparición y extinción de géneros , especies y poblaciones.
La hipótesis del origen único
Esta hipótesis ya fue propuesta a principios de siglo a partir de los hallazgos de fósiles humanos que se produjeron en Europa y Asia, que tenían como protagonistas principales a los neandertales y a los hombres de Cromañón. Después del estudio del neandertal de la Chapelle-aux-Saints, el paleontólogo francés Marcellin Boule propuso que los neandertales no eran nuestros antepasados, sino que Homo sapiens había evolucionado en alguna otra parte del mundo, y que, posteriormente, había colonizado Europa provocando la extinción de los neandertales. Los prehistoriadores observaban que en las secuencias estratigráficas de los yacimientos europeos los neandertales siempre aparecían en niveles por debajo (y por lo tanto más antiguos) a los de los hombres de Cromañón. Además, los primeros estaban asociados a industrias líticas del Paleolítico medio , mientras que los segundos aparecían junto a industria lítica del Paleolítico superior . El cambio del tipo de humanos y del tipo de industria lítica aparecía en el registro arqueológico de forma brusca.
Los fósiles humanos del Pleistoceno medio y superior, el periodo clave para entender el proceso de aparición de Homo sapiens,se han multiplicado en la segunda mitad del siglo XX. A partir de éstos, numerosos paleoantropólogos, entre los que destacan Gunter Bräuer y Christopher Stringer, opinan que los humanos modernos aparecieron en África hace entre 300.000 y 100.000 años, y a partir de este continente emigraron al resto del Viejo Mundo, reemplazando a las distintas humanidades presentes en Asia y Europa. En este proceso colonizador se extinguieron los neandertales europeos y los Homo erectus asiáticos, que habían aparecido como resultado de evoluciones locales en condiciones de aislamiento genético. Esta hipótesis, del origen exclusivamente africano, fue bautizada "Out of Africa" o "Fuera de África". Por lo tanto, en la evolución humana se habrían producido un mínimo de dos salidas de África: una primera, durante el Pleistoceno inferior, protagonizada por los antepasados de los primeros pobladores de Asia (Homo erectus) y Europa (Homo antecessor), los cuales encontraron ambos continentes en estado virgen; y una segunda salida más reciente, en el Pleistoceno superior, protagonizada por Homo sapiens que avanzó sobre territorios que ya estaban ocupados por otros humanos y que con el tiempo acabó sustituyéndolos.
La evidencia fósil
Uno de los principales problemas en el estudio de los primeros hombres modernos es de tipo metodológico, ya que el rango de edades de los yacimientos de interés queda fuera de la técnicas de datación tradicionales. Las técnicas radiocarbónicas no pueden datar materiales de más de 30.000 o 40.000 años y, en muy pocos casos, se dispone de materiales de tipo volcánico para aplicar otras técnicas, como el K-Ar o el Ar-Ar. A partir de los años ochenta, esta dificultad ha ido desapareciendo gracias al desarrollo de nuevas técnicas de datación: las series de Uranio, la termoluminiscencia y la resonancia de espín electrónico. La aplicación de estas técnicas ha confirmado que el hombre moderno estaba presente en África cuando los neandertales todavía vivían en Europa y Asia occidental, y el Homo erectus todavía poblaba la isla de Java.
Primeras investigaciones en el Próximo Oriente
La investigación sobre los homínidos del Próximo Oriente comenzó en 1929, cuando la arqueóloga Dorothy Garrod inició un proyecto quinquenal de excavación en las cuevas de Skhul, Tabun y el-Wad en el Monte Carmelo (Israel), a la que se unió más tarde Theodore McCown. Los dos primeros yacimientos proporcionaron varios esqueletos humanos parciales, resultado de posibles enterramientos. Inicialmente, se propusieron diversas hipótesis sobre cuales eran las poblaciones representadas en ambas cuevas:  eran formas intermedias entre los neandertales y los humanos modernos. posibles híbridos entre ambas poblaciones. representaban una población que evolucionó localmente.
eran formas intermedias entre los neandertales y los humanos modernos. posibles híbridos entre ambas poblaciones. representaban una población que evolucionó localmente.
Entre 1935 y 1975 se excavó una muestra mayor de humanos modernos de tipo primitivo en Jebel Qafzeh, cerca de Nazaret, y se descubrieron más neandertales en las cuevas de Kebara, también en el Monte Carmelo, y en Amud, junto al mar de Galilea. Finalmente, gracias a las aportaciones de los paleoantropólogos Clark Howell y Bernard Vandermeersch, la conclusión que se aceptó fue que Tabun, Kebara y Amud habían sido habitados por los neandertales. En cambio, los fósiles de Skhul y Qafzeh pertenecían a humanos modernos con rasgos primitivos (algunos presentan torus supraorbitarios muy marcados). Estos humanos tenían un cráneo con una gran capacidad craneal, forma globosa, con la frente alta, nariz estrecha y las mandíbulas poseían mentón. Su esqueleto postcraneal era menos robusto que los neandertales y el tronco era más estrecho y esbelto. La estructura de su esqueleto es muy similar a la que poseen los humanos actuales de tipo tropical, muy altos y con miembros largos. Es decir, su morfología es muy similar a la de la humanidad actual y los fósiles humanos de Skhul y Qafzeh pertenecen a nuestra misma especie : Homo sapiens.
Nuevas dataciones e interpretaciones en el Próximo Oriente Así, por los años setenta, la opinión general era que en Oriente Medio y en Europa los humanos modernos llegaron después de los neandertales y, en consecuencia, podían haber descendido de ellos. Tanto los neandertales como los humanos modernos aparecían asociados al mismo tipo de tecnología lítica del Modo 3 o Paleolítico medio : el Musteriense . Pero esta reconstrucción cronológica chocaba con los análisis de la fauna fósil de Tabun y Qafzeh realizados por E. Tchernov y con los estudios de la secuencia polínica. En algunos niveles de Qafzeh, que contenían fósiles humanos, Tchernov había hallado varias especies de roedores arcaicos que no estaban representadas en el nivel C de Tabun (de donde proceden el esqueleto de neandertal y una mandíbula aislada posiblemente de Homo sapiens). La aplicación de las nuevas técnicas de datación a los fósiles de Skhul y Qafzeh vino a confirmar la sospecha de que ambos yacimientos eran más antiguos de lo que se pensaba. Los fósiles humanos de Skhul y Jebel Qafzeh han sido datados por termoluminiscencia, resonancia de espín electrónico y series de uranio, dando una antigüedad en torno a los 100.000 años. El esqueleto neandertal de Tabun posiblemente tiene en torno a los 110.000 años, pero en cambio los fósiles de neandertales de Kebara, Amud y Dederiyeh son más recientes, entre los 85.000 y los 50.000 años.
Estos resultados refutaron la relación genealógica defendida durante mucho tiempo y los neandertales, que mostraban aquellos esqueletos robustos, no podían ser los antepasados de Homo sapiens, con esqueletos más gráciles. Tampoco era posible seguir vinculando la existencia de los humanos modernos a la revolución tecnológica del Paleolítico superior , porque cuando elHomo sapiens aparece en escena en el Próximo Oriente, su modo de vida y su tipo de industria lítica era indistinguible del de los neandertales. Los humanos modernos estaban presentes en el Próximo oriente hace 100.000 años. De hecho, puede que tengan una genealogía local bastante amplia. En la cueva de Zuttiyeh, cerca de Amud, Turville-Petre encontró en 1925 un fragmento de cráneo (principalmente la región frontal) que podría tener entre los 200.000 y los 250.000 años de antigüedad. Este individuo, aunque muy fragmentario, podría haber formado parte de la población de la que descendieron los pobladores de Skhul y Qafzeh. En cambio, los neandertales no parece que hayan tenido antepasados en el Próximo Oriente y por lo tanto son descendientes de los neandertales europeos. En primer lugar, como atestiguaría el esqueleto de Tabun con una antigüedad de 110.000-100.00 años, el Próximo Oriente fue ocupado por los neandertales, pero los humanos modernos los sustituyeron hace unos 100.000 años. Tenemos que destacar que en este yacimiento, en el mismo nivel estratigráfico (nivel C), apareció un esqueleto claramente de un neandertal, y una mandíbula que parece pertenecer a un humano moderno. Pero, existe la posibilidad de que el esqueleto fuese de edad posterior y que hubiese sido introducido en el nivel C al enterrar el cuerpo. Posteriormente, entre hace 60.000 y 40.000 años, los neandertales desplazaron a los humanos modernos hacia el sur y, finalmente, hace 40.000 años, los neandertales desaparecieron del Próximo Oriente. Según el arqueólogo israelí Ofer Bar-Yosef, los neandertales habrían poblado el Próximo Oriente en los momentos de fríos intensos, que habrían obligado a los neandertales a desplazarse hacia el sur. La desaparición de los neandertales coincide con la aparición de las industrias líticas del Paleolítico superior en el Próximo Oriente y en Europa.
Los fósiles africanos En torno a un millón de años de antigüedad hay un gran vacío de fósiles humanos en África, en donde podría encontrarse una población de características muy similares a la encontrada en la Gran Dolina de la Sierra de Atapuerca, y que ha sido asignada a la nueva especie Homo antecessor. A partir de este antepasado común, en Europa las poblaciones humanas evolucionaron dando lugar a los neandertales, en África evolucionaron hacia nuestra especie. Los fósiles africanos del Pleistoceno medio son muy similares morfológicamente a los humanos mesopleistocenos europeos. Aunque la mayor parte de las dataciones son dudosas, en este grupo podemos situar a los fósiles de Bodo, Eyasi, Ndutu, Salé y Broken Hill. Merced a esta similitud, muchos científicos incluyen a estos fósiles africanos junto a los fósiles del Pleistoceno medio europeo en la especie Homo heidelbergensis. Pero la similitud morfológica es únicamente debido a su estrecha relación evolutiva, porque comparten un antepasado común muy próximo en el tiempo. En los fósiles de la línea evolutiva africana pueden observarse algunas características que darán lugar a Homo sapiens, y por lo tanto deben clasificarse como una especie distinta:Homo rhodesiensis.
Entre hace 200.000 años y 100.000 años, en los fósiles humanos hallados en África, empiezan a aparecer características propias de la humanidad actual. Es el caso de los fósiles procedentes de los niveles superiores de Laetoli (cráneo L.H. 18), de las formaciones superiores del valle del río Omo (cráneo Omo Kibish 2) y de otros restos sudafricanos (Border Cave, Florisbad, Cave of Hearths). Estos fósiles presentan una prognatismo facial más reducido, mayores capacidades craneales y una frente más elevada y convexa. En el norte de África, también se han encontrado fósiles que pueden representar a estas poblaciones transicionales que evolucionaban hacia Homo sapiens. En Jebel Irhoud, en Marruecos, se han encontrado restos de unos cinco individuos de entre 100.000 y 200.000 años de antigüedad. En el cráneo más completo, Jebel Irhoud 1, puede observarse un neurocráneo primitivo, con una gran capacidad craneal (1305 cc), unido a una cara de morfología muy moderna.
Estos fósiles del Pleistoceno medio final y Pleistoceno superior de África podrían ser calificados de premodernos. Los pocos cráneos que las conservan, tienen caras gráciles similares a las de Homo sapiens, tienen mayores capacidades craneales y sus neurocráneos son menos robustos y más redondeados. Sin embargo, en ellos no se observa todavía el conjunto de características de un cráneo moderno, casi esférico, con la bóveda alta y la frente vertical. Uno de los fósiles más antiguos de Homo sapiens en África podía ser el esqueleto parcial de Omo-Kibish 1 (Etiopía), hallado por Richard Leakey en 1967, con una antigüedad entre 50.000 y 130.000 años. Tanto su cráneo, muy fragmentario, como su esqueleto postcraneal presenta una morfología muy moderna, más esbelta y grácil que esqueletos primitivos. En el complejo de cuevas de Klasies River Mouth, en Sudáfrica, se han encontrado fósiles humanos muy fragmentarios con una edad entre 90.000 y 120.000 años, pero que son anatómicamente muy similares a los de la humanidad actual. Estos restos de Klasies River Mouth están quemados y presentan numerosas marcas de corte en su superficie porque fueron consumidos por otros humanos.
La expansión de los humanos modernos Si en África se observa una transición entre los fósiles más arcaicos hasta fósiles que pertenecen a nuestra especie, en otras partes del mundo la sustitución entre los homínidos más primitivos y los Homo sapiens fue mucho más abrupta. Como atestiguan los fósiles de Qafzeh y Skhul, los humanos modernos ya estaban en el Próximo Oriente hace 100.000 años, aunque luego fueron desplazados de allí por los neandertales. En un primer momento los neandertales ganaron la partida a los Homo sapiens, pero después la perdieron de forma definitiva. En Europa se observa una sustitución brusca de la industria Musteriense de Modo 3 por el Auriñaciense, la primera industria delPaleolítico superior o Modo 4. El Musteriense se atribuye a los neandertales y el Auriñaciense lo traerían consigo los Homo sapiens (la confrontación entre neandertales y cromañones fue un proceso más complejo que veremos con detalle en la siguiente Unidad didáctica). El Homo sapiens se expandió por Europa hace unos 40.000 años y lo encontramos en numerosos yacimientos posteriores a esa fecha. Los humanos modernos también se expandieron hasta el extremo oriental de Eurasia, llegando a Australia hace entre 60.000 y 50.000 años, fechas ligeramente más antiguas que el poblamiento de Europa. En los yacimientos de Australia únicamente se encuentran utensilios líticos de Modo 3 y no de Modo 4, tal y como aparecen en Europa asociados a Homo sapiens. Es muy posible, que el poblamiento de Asia y Australia sea más antiguo que el de Europa y fuese anterior a la "invención" del Modo 4, realizada por un grupo de Homo sapiens, en un lugar aún por determinar (en el Próximo oriente o en África). Fue este desarrollo tecnológico y su mayor complejidad cultural lo que dió a los humanos modernos algún tipo de ventaja sobre los neandertales, causando el éxito evolutivo de los primeros.
La evidencia molecular - El código genético
La molécula responsable de la herencia biológica es el ácido desoxirribonucleico (ADN), que lleva codificada en su estructura química la información que constituye a cada ser vivo. El ADN está organizado en cromosomas, en pares homólogos, dentro del núcleo celular. El número de cromosomas varía de una especie a otra (por ejemplo, los humanos tenemos 23 pares de cromosomas, los chimpancés, orangutanes y gorilas 24 pares, los macacos 21 pares). En la reproducción sexual, se produce una división celular especial, llamada meiosis , por la que cada una de las células reproductoras (óvulos y espermatozoides) contiene sólo la mitad de los cromosomas. En el interior de cada una de nuestras células hay un núcleo que contiene la mayor parte de nuestro ADN. Pero fuera del núcleo se encuentra el ADN mitocondrial (ADNmt): numerosas copias de un filamento circular de ADN, constituidos por unos 16.000 pares de bases (en comparación con los tres mil millones de pares que constituyen el ADN nuclear). Las mitocondrias son pequeños orgánulos responsables del metabolismo energético de las células (al igual que los cloroplastos de las plantas que capturan la energía solar) y que también poseen su propio genoma . El ADNmt es muy interesante porque es relativamente pequeño, presenta una tasa muy elevada de mutaciones, y tiene la peculiaridad que sólo se hereda por línea materna. A diferencia del ADN nuclear, que heredamos la mitad de nuestra madre y la mitad de nuestro padre, el ADNmt es transmitido únicamente por las hembras, porque los espermatozoides sólo transportan ADNmt en su cola y se pierde en el proceso de fecundación. Debido a este modo de herencia exclusiva materna, no existe recombinación en el ADNmt entre los genes maternos y paternos, un hecho que puede enmascarar la historia evolutiva del genoma y hace mucho más difícil su investigación.
La diversidad genética actual
A partir de los años setenta, diferentes laboratorios del mundo empezaron a investigar la diversidad genética de la humanidad actual utilizando el ADNmt. Una de las primeras observaciones fue la escasa variación genética que existe entre los humanos, aproximadamente sólo el 10% de la variabilidad genética de la observada en los chimpancés. Esta baja variabilidad genética, que también se observa en el ADN nuclear y en otros marcadores genéticos implica un origen muy reciente de la humanidad actual. En 1983, el equipo de Douglas Wallace publicó el primer estudio de ADNmt en la humanidad actual llegando a varias conclusiones que, con ligeras modificaciones, siguen siendo válidas hoy en día:  1) la variación en el ADNmt de los humanos modernos es muy pequeña y implica que el origen de nuestra especie es muy reciente, y se sitúa aproximadamente en hace 200.000 años. 2) De todas las poblaciones actuales estudiadas, entre las africanas se da la mayor variabilidad. Si la tasa de mutación es igual en todas las poblaciones, este dato sugiere que el linaje humano es más antiguo en África y su origen debe situarse en este continente. Inicialmente Wallace y sus colaboradores pensaron que esta mayor variabilidad se debía a una mayor tasa de mutación en las poblaciones africanas, y que el origen de nuestra especie se encontraba en Asia, pero muy pronto cambiaron su opinión defendiendo a África como cuna de la humanidad. Uno de los estudios más importantes fue el publicado por R. Cann, M. Stoneking y A. Willson en 1987, que analizaron el ADNmt de 147 individuos procedentes de distintas poblaciones geográficas (ver Figura 6.2.9). Estos autores midieron la diferencias de una parte muy importante de la secuencia del ADN mitocondrial en cada uno de los individuos y elaboraron un árbol genealógico utilizando el principio de la parsimonia (que trata de encontrar el árbol que contiene el menor número de cambios o de pasos evolutivos).
1) la variación en el ADNmt de los humanos modernos es muy pequeña y implica que el origen de nuestra especie es muy reciente, y se sitúa aproximadamente en hace 200.000 años. 2) De todas las poblaciones actuales estudiadas, entre las africanas se da la mayor variabilidad. Si la tasa de mutación es igual en todas las poblaciones, este dato sugiere que el linaje humano es más antiguo en África y su origen debe situarse en este continente. Inicialmente Wallace y sus colaboradores pensaron que esta mayor variabilidad se debía a una mayor tasa de mutación en las poblaciones africanas, y que el origen de nuestra especie se encontraba en Asia, pero muy pronto cambiaron su opinión defendiendo a África como cuna de la humanidad. Uno de los estudios más importantes fue el publicado por R. Cann, M. Stoneking y A. Willson en 1987, que analizaron el ADNmt de 147 individuos procedentes de distintas poblaciones geográficas (ver Figura 6.2.9). Estos autores midieron la diferencias de una parte muy importante de la secuencia del ADN mitocondrial en cada uno de los individuos y elaboraron un árbol genealógico utilizando el principio de la parsimonia (que trata de encontrar el árbol que contiene el menor número de cambios o de pasos evolutivos).
1) la variación en el ADNmt de los humanos modernos es muy pequeña y implica que el origen de nuestra especie es muy reciente, y se sitúa aproximadamente en hace 200.000 años. 2) De todas las poblaciones actuales estudiadas, entre las africanas se da la mayor variabilidad. Si la tasa de mutación es igual en todas las poblaciones, este dato sugiere que el linaje humano es más antiguo en África y su origen debe situarse en este continente. Inicialmente Wallace y sus colaboradores pensaron que esta mayor variabilidad se debía a una mayor tasa de mutación en las poblaciones africanas, y que el origen de nuestra especie se encontraba en Asia, pero muy pronto cambiaron su opinión defendiendo a África como cuna de la humanidad. Uno de los estudios más importantes fue el publicado por R. Cann, M. Stoneking y A. Willson en 1987, que analizaron el ADNmt de 147 individuos procedentes de distintas poblaciones geográficas (ver Figura 6.2.9). Estos autores midieron la diferencias de una parte muy importante de la secuencia del ADN mitocondrial en cada uno de los individuos y elaboraron un árbol genealógico utilizando el principio de la parsimonia (que trata de encontrar el árbol que contiene el menor número de cambios o de pasos evolutivos).
Los resultados coincidían con los estudios anteriores y pueden resumirse en dos puntos:
2) El grupo africano presenta una variabilidad del ADNmt relativamente mayor. Es decir, el grupo africano es el más antiguo de todos porque ha acumulado mayores diferencias, y este grado de variabilidad puede usarse para medir su antigüedad. La tasa promedio de mutación del ADNmt fue calculada midiendo el grado de divergencia genética acumulada por las poblaciones de Papúa Nueva Guinea, colonizada hace unos 40.000 años. El valor de la divergencia entre dos líneas era del 2-4 % por cada millón de años, es decir de 2 a 4 posiciones de nucleótidos por cada 100 posiciones cambiarán respecto a dosgenomas . Esta tasa de mutación ha sido confirmada por otros laboratorios con datos procedentes de humanos y de otrasespecies de mamíferos. Teniendo en cuenta la tasa de mutación del ADNmt, Cann y sus colaboradores concluían que todos los humanos tenemos una antepasada común que vivió en África hace unos 200.000 años, la denominada "Eva africana" o " Eva mitocondrial ".
Los "relojes moleculares"
Estos análisis genéticos recibieron muchas críticas, principalmente por la forma en que estimaron el tiempo de divergencia. El establecimiento de estos "relojes moleculares" es el mayor problema de los estudios genéticos sobre el origen de la humanidad, ya que la única forma de establecerlos es utilizando el registro fósil. También es muy difícil cuantificar la influencia que pueden tener los "cuellos de botella" evolutivos (reducciones en el tamaño de la población antepasada). La escasa diversidad genética que muestran las poblaciones humanas de fuera de África son consecuencia de que derivan de una pequeña fracción de la población africana original, lo que se denomina el efecto fundador. Sin embargo, existen diferentes opiniones en cuanto al tamaño de esos colonizadores que influye en el cálculo del tiempo de divergencia de los linajes evolutivos. También se ha cuestionado el hecho de que una mayor diversidad genética de las poblaciones africanas implique una mayor antigüedad de la humanidad, sino que esto puede ser debido al mayor tamaño de la población ancestral. Para contrastar los resultados del estudio del ADNmt se analizó una parte del ADN nuclear que se transmite sólo por vía paterna, y se localiza en el cromosoma Y. Se estudiaron los polimorfismos del cromosoma Y, los cuales pueden resumirse en unos pocos tipos (haplotipos). Los resultados coinciden con los del ADNmt: la humanidad moderna tuvo un antepasado varón que vivió en África hace entre 100.000 y 200.000 años. También, Luca Cavalli-Sforza y sus colaboradores analizando otros segmentos de otros cromosomas del ADN nuclear llegaron a conclusiones similares situando el origen del hombre moderno hace unos 120.000 en África, y además trazaba la ruta que siguió la colonización del Viejo continente: en primer lugar se produjo la separación de los linajes africanos y asiáticos hace unos 50.000-60.000 años, y posteriormente se separaron los europeos hace 30.000-40.000 años.
Estado actual de los estudios genéticos La discusión científica continúa en la actualidad, abarcando desde problemas de muestreo, metodológicos, estadísticos, etc. Además, en los últimos años han continuado apareciendo estudios de diferentes equipos que han utilizado diversos tipos de información genética (cromosoma Y, marcadores genéticos, polimorfismos, microsatélites, complejo principal de histocompatibilidad, etc.). Resumiendo, la mayor parte de estudios genéticos confirman la hipótesis de "Out of Africa", establecida a partir de los fósiles, sobre el origen africano y reciente de nuestra especie . Es decir, el origen de los humanos modernos es muy reciente porque la variación del ADN es muy limitada. En segundo lugar, las poblaciones africanas exhiben mayor variabilidad debido a la mayor longitud de su línea evolutiva. En cambio, no se encuentra la amplia variación genética que predice la hipótesis evolutiva multirregional, que implicaría un origen antiguo de la humanidad y en la que ninguna población debería presentar un grado significativamente mayor de variación que otra.
Genes fósiles
Como hemos visto más arriba, la Genética aborda los problemas evolutivos a partir del estudio de la variabilidad genética de seres vivos. Sólo en casos excepcionales puede estudiarse directamente los genes de especies del pasado. La molécula de ADN se oxida con bastante rapidez tras la muerte del organismo y es muy difícil evitar su deterioro. El ADN fósil más antiguo que se había podido recuperar procedía de los mamuts conservados en el suelo helado de Siberia, con unos 50.000 años de antigüedad. Pero recientemente, el equipo de Svante Pääbo ha conseguido aislar pequeños fragmentos de ADN de un neandertal. Con todas las precauciones necesarias para evitar la contaminación con ADN actual, tomaron una muestra de varios gramos del interior del húmero en el esqueleto de Neander, con 60.000 años de antigüedad. La muestra fue analizada por dos laboratorios distintos para poder contrastar los resultados y comprobar la inexistencia de contaminación. El ADN mitocondrial que encontraron estaba muy fragmentado pero, uniendo la secuencia de numerosos fragmentos distintos que se solapaban, ha sido posible reconstruir dos segmentos de ADNmt: un segmento de 333 pares de bases , que corresponde con la región hipervariable I de la región de control del ADNmt; y un segundo fragmento de 340 pares de bases de la región hipervariable II de la región de control del ANDmt. La comparación de estos dos fragmentos de secuencia de ADNmt con la secuencia de ADNmt de 9 chimpancés (7 Pan troglodytes y 2 Pan paniscus) y con 663 tipos de ADNmt de distintas poblaciones humanas actuales arrojó los siguientes resultados:  1) Los humanos actuales y los chimpancés difieren en promedio en 93 posiciones de las secuencias de bases . 2) Entre las poblaciones actuales la diferencia promedio es de 11 posiciones. 3) La secuencia fósil de neandertal tiene una diferencia media respecto de los humanos actuales en 35 posiciones. 4) Las menores diferencias se dan entre la secuencia neandertal y las poblaciones africanas actuales. La primera conclusión de este equipo es que el ADNmt analizado no procedía de ninguna contaminación, sino que era realmente ADNmt de un neandertal. Además, las diferencias en la secuencia de ADNmt ha llevado a estos autores a concluir que: 1) la separación entre las líneas evolutivas de los neandertales y de los humanos modernos se produjo hace 465.000 (con límites de confianza al 95% de 317.000 y 741.000 años); y 2) que el origen de la diversidad humana actual se produjo en el continente africano hace alrededor de 163.000 años (límites de confianza de 111.000 y 260.000 años). La primera de las fechas, que indica la separación de la línea de los neandertales, está situada en más de medio millón de años, y es compatible con los resultados del equipo investigador de la Sierra de Atapuerca. Éstos proponen que los fósiles humanos hallados en el nivel TD6 de la Gran Dolina, con 800.000 años de antigüedad, representan la especie antecesora de los neandertales y de los humanos modernos, que se denomina Homo antecessor. La segunda de las fechas, que sitúa el origen del hombre moderno en hace más de 100.000 años, confirma los estudios anteriores del ANDmt de los humanos actuales. Otra cuestión que puede abordarse es la posibilidad de si los neandertales y los humanos modernos hubieran hibridado. A partir del ANDmt de neandertal no puede excluirse completamente, pero ambos grupos han evolucionado genéticamente de forma aislada durante un sustancial periodo de tiempo, y para estos autores los neandertales no han contribuido al acervo genético de ADNmt de la humanidad actual.
1) Los humanos actuales y los chimpancés difieren en promedio en 93 posiciones de las secuencias de bases . 2) Entre las poblaciones actuales la diferencia promedio es de 11 posiciones. 3) La secuencia fósil de neandertal tiene una diferencia media respecto de los humanos actuales en 35 posiciones. 4) Las menores diferencias se dan entre la secuencia neandertal y las poblaciones africanas actuales. La primera conclusión de este equipo es que el ADNmt analizado no procedía de ninguna contaminación, sino que era realmente ADNmt de un neandertal. Además, las diferencias en la secuencia de ADNmt ha llevado a estos autores a concluir que: 1) la separación entre las líneas evolutivas de los neandertales y de los humanos modernos se produjo hace 465.000 (con límites de confianza al 95% de 317.000 y 741.000 años); y 2) que el origen de la diversidad humana actual se produjo en el continente africano hace alrededor de 163.000 años (límites de confianza de 111.000 y 260.000 años). La primera de las fechas, que indica la separación de la línea de los neandertales, está situada en más de medio millón de años, y es compatible con los resultados del equipo investigador de la Sierra de Atapuerca. Éstos proponen que los fósiles humanos hallados en el nivel TD6 de la Gran Dolina, con 800.000 años de antigüedad, representan la especie antecesora de los neandertales y de los humanos modernos, que se denomina Homo antecessor. La segunda de las fechas, que sitúa el origen del hombre moderno en hace más de 100.000 años, confirma los estudios anteriores del ANDmt de los humanos actuales. Otra cuestión que puede abordarse es la posibilidad de si los neandertales y los humanos modernos hubieran hibridado. A partir del ANDmt de neandertal no puede excluirse completamente, pero ambos grupos han evolucionado genéticamente de forma aislada durante un sustancial periodo de tiempo, y para estos autores los neandertales no han contribuido al acervo genético de ADNmt de la humanidad actual.
La evidencia genética vs. la evidencia fósil En muchos ocasiones se presenta la evidencia genética como prueba definitiva frente a la incertidumbre del registro fósil y de sus interpretaciones subjetivas. Dos han sido los focos de discusión entre paleontólogos y genéticos: el primero para situar la divergencia entre el linaje que conduce a los humanos y el que conduce a los chimpancés; y en segundo lugar el origen geográfico y temporal de la humanidad actual. En el primer caso, se llegó a un consenso intermedio, pero en el segundo caso los estudios genéticos sólo han confirmado la hipótesis que había sido propuesta a partir de los fósiles. Los estudios genéticos han aportado muchas pruebas sobre el cómo y el dónde del origen de nuestra especie , pero sus conclusiones sobre el cuándo son más cuestionables. Uno de los principales problemas de los estudios genéticos es la situación temporal de las divergencias. Para establecer los "relojes moleculares" se deben tomar previamente algunas presupuestos: las semejanzas genéticas entre individuos han de ser función de su parentesco, y directamente proporcionales al tiempo de divergencia desde un antepasado común; la tasa de mutación ha de ser constante; y los genes estudiados deben ser neutros y que no actúe sobre ellos la selección natural. Además, los genetistas deben utilizar el registro fósil para calibrar las divergencias que hallan. En definitiva, los datos paleontológicos y genéticos deben apoyarse y no son independientes. Hoy en día, son muy pocos los científicos que sostienen la hipótesis multirregional del origen de los humanos modernos. Y éstos utilizan las críticas y revisiones que reciben los estudios moleculares para socavar los cimientos de la hipótesis "Out of Africa", establecida a partir de los fósiles. Los análisis genéticos han reforzado la opinión de muchos paleontólogos de que los neandertales no son antepasados de la humanidad actual, sino que ambos grupos comparten un antepasado común lejano en el tiempo.

.jpg)
0 comentarios: